Promotional Features


Finding Bioglena™ - the optimal ingredient for immune formulation
β-glucan polysaccharides are touted as nature’s immune-modulator, helping to prime immune cells, supporting and balancing optimal natural immunity. It helps the immune system to respond efficiently when pathogens attack.
A balanced immune system is essential for our survival, function and quality of life. When the immune system is imbalanced it may either be over activated causing autoimmune diseases or allergic reactions or deficient leading to higher susceptibility to pathogens and infectious diseases.
The immune system has evolved to recognize β-glucans as they are found on the surface of disease-causing fungi and are specifically recognized by cellular receptors which activate the immune system. These receptors are mainly present in innate immune cells, for example macrophages. The primary receptor for recognizing β-glucans is Dectin-1, although it is not the only one (1,2)
Trained immunity, a newly identified immune mechanism, describes a functional reprogramming of innate immune cells following exposure to stimuli, which leads to an enhanced response toward a second stimulus. In other words, trained immunity is the ability of the innate immune system to recall or adapt to a first challenge to respond more rapidly and robustly to a secondary challenge by a similar or dissimilar microbial or fungi stimulus (3,4,5). β-glucans are typical inducers of trained immunity (6).
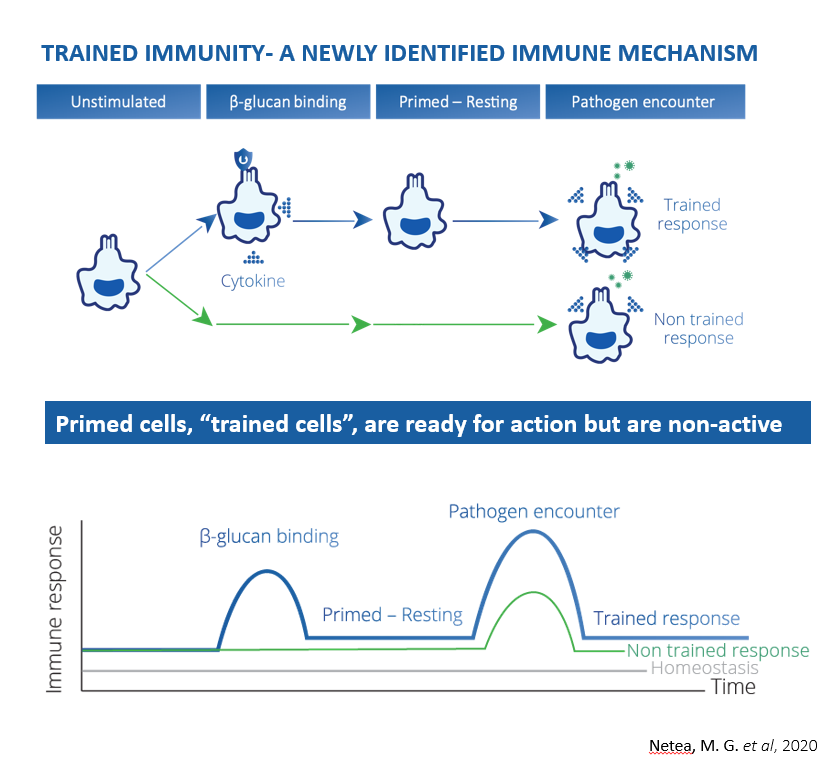
ß-glucans induce trained immunity
β-glucans are prototypical inducers of trained immunity (6), which involves three phases sequentially:
1) First, unstimulated immune cells, such as macrophages, bind β-glucan via the dectin-1 receptor and are thereby activated and undergo a functional epigenetic reprogramming and associated metabolic changes, preparing the cells for future encounters with pathogens.
2) In the following stage the trained and primed cells are in a resting and non-active mode.
3) The resting period ends when trained/primed cells encounter a pathogen, leading to their response which is more effective and powerful than the response of non-trained/non-primed cells (4,5).
Euglena gracilis a high source of Beta glucans
Euglena gracilis (E. grcaillis ) is a single-cell microalga, living in most freshwater habitats.
Euglena gracilis naturally produces linear 1-3 β-glucans called paramylon, which are stored as free-floating granules in the cell and are known for their immuno-modulatory functions.
E. grcaillis uses the β-glucans as a principal energy storage compound and can accumulate a large amount of β-glucans within the cell. The algae also contain complete proteins, essential vitamins, and minerals.
Paramylon (linear 1-3 β-glucans) has been shown to bind to the dectin-1 receptor, leading to activation of immune cells, aimed at pathogen elimination, including trained immunity (4).
BioGlena™ - Solabia-Algatech Nutrition's high source of the Beta-glucan
BioGlena™️ is a 100% natural whole Euglena gracilis microalgae product, produced by a proprietary fermentation technology by Solabia-Algatech Nutrition, for high β-glucan content.
BioGlena contains a minimum of 55% linear 1,3 β-glucan, complete protein, essential vitamins and minerals.
BioGlena™️ is easily digested and the β-Glucan is easily released from the whole algae in the stomach and is then picked up by immune cells in the intestine and transported to key immune sites in the body. This initiates a cascade of immune signaling and immune cell priming, ultimately strengthening our natural innate and adaptive immune support against pathogens.
BioGlena™️ requires no extraction procedure, hence contains no additives and solvents, and is suitable and safe for the whole family for daily use.
BioGlena™ has been recognized by the FDA as a G.R.A.S.
References:
1) Brown, G. D., Herre, J., Williams, D. L., Willment, J. A., Marshall, A. S. J., & Gordon, S. (2003). Dectin-1 mediates the biological effects of β-glucans. Journal of Experimental Medicine, 197(9), 1119–1124. https://doi.org/10.1084/jem.20021890
2) Brown, G. D., Taylor, P. R., Reid, D. M., Willment, J. A., Williams, D. L., Martinez-Pomares, L., Wong, S. Y. C., & Gordon, S. (2002). Dectin-1 is a major β-glucan receptor on macrophages. Journal of Experimental Medicine, 196(3), 407–412. https://doi.org/10.1084/jem.20020470
3) Netea, M. G., Domínguez-Andrés, J., Barreiro, L. B., Chavakis, T., Divangahi, M., Fuchs, E., Joosten, L. A. B., van der Meer, J. W. M., Mhlanga, M. M., Mulder, W. J. M., Riksen, N. P., Schlitzer, A., Schultze, J. L., Stabell Benn, C., Sun, J. C., Xavier, R. J., & Latz, E. (2020). Defining trained immunity and its role in health and disease. Nature Reviews Immunology, 20(6), 375–388. https://doi.org/10.1038/s41577-020-0285-6
4) Paris, S., Chapat, L., Martin-Cagnon, N., Durand, P. Y., Piney, L., Cariou, C., Bergamo, P., Bonnet, J. M., Poulet, H., Freyburger, L., & De Luca, K. (2020). β-Glucan as Trained Immunity-Based Adjuvants for Rabies Vaccines in Dogs. Frontiers in Immunology, 11(October), 1–14. https://doi.org/10.3389/fimmu.2020.564497
5) Bekkering, S., Blok, B. A., Joosten, L. A. B., Riksen, N. P., Van Crevel, R., & Netea, M. G. (2016). In Vitro experimental model of trained innate immunity in human primary monocytes. Clinical and Vaccine Immunology, 23(12), 926–933. https://doi.org/10.1128/CVI.00349-16
6) Quintin, J., Saeed, S., Martens, J. H. A., Giamarellos-Bourboulis, E. J., Ifrim, D. C., Logie, C., Jacobs, L., Jansen, T., Kullberg, B. J., Wijmenga, C., Joosten, L. A. B., Xavier, R. J., Van Der Meer, J. W. M., Stunnenberg, H. G., & Netea, M. G. (2012). Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host and Microbe, 12(2), 223–232. https://doi.org/10.1016/j.chom.2012.06.006
